Succinate Induces Liver Damage and Hepatic Fibrosis in a Mouse Model
Article information
Abstract
Hepatic stellate cells (HSCs) play a key role in liver fibrosis. Succinate and succinate receptor (GPR91) signaling pathway are involved in the activation, proliferation, and migration of HSCs. We investigated whether succinate may induce hepatic fibrosis. The mice were randomly divided into 2 groups —the control group (chow diet-fed mice, n = 26) and sodium succinate group (2% sodium succinate + chow diet, n = 38). Each diet was provided for 16 weeks. Mice administered an oral diet of 2% sodium succinate for sixteen weeks lost body weight and had increased serum alanine transaminase and hepatic triglyceride contents compared to those in the control mice. Moreover, mice fed with sodium succinate showed increased expression of the alpha smooth muscle actin protein and gene in the liver at 8 weeks of feeding and increased fibrosis in their histology at 16 weeks of feeding. However, the expression of the GPR91 protein and mRNA increased at 4 weeks of feeding, but decreased at 8 and 16 weeks of feeding. These results suggest that an oral succinate diet could induce liver damage and liver fibrosis in mice and that GPR91 signaling might be an early marker or sensor of hepatic fibrosis development.
Introduction
Liver fibrosis is a common pathological condition that occurs because of chronic liver injury resulting from various etiological factors such as viruses, toxins, alcohol [1]. Succinate is an intermediate in the tricarboxylic acid (TCA) cycle that plays an important role in the generation of Adenosine triphosphate in the mitochondria. Different reports have shown that succinate could regulate various cell functions related to epigenetics, tumor genesis, inflammation, signal transduction, and paracrine modulation in various tissues, including the liver, kidney, heart, retina, and immune tissues [2,3,4]. Our previous study revealed that succinate increased fibrogenic markers, inflammatory cytokine expression, and the proliferation and migration of LX-2 cells, and that administration of intraperitoneal succinate in mice showed increased alpha smooth muscle actin (α-SMA) and inflammatory cytokine protein expression, apart from the histological changes [5]. A recent study demonstrated that administration of succinate with drinking water could control the activation of adipose tissue thermogenesis [6]. Although the effect of succinate on the regulation of hepatic stellate cells (HSCs) has been demonstrated, the effect of succinate on liver fibrosis and damage in mice in a time-dependent manner is not known. Further, a new mouse model for liver fibrosis using succinate is still lacking. Therefore, we conducted this research to validate whether succinate induces liver fibrosis in a mouse model.
Materials and Methods
Animal procedures
Male C57BJ6 mice weighing 22-25 g and aged 6 weeks were provided by Doo Yeol Biotech (Seoul, Korea). All mice were reared at 22 ± 1°C with free access to food and water and a 12 h light/dark cycle. All animal experiments were approved by the Institutional Animal Care and Use Committee of Kangwon National University (Protocol approval # KW-180103-1). The mice were randomly divided into 2 groups —the control group (chow diet-fed mice, n = 26) and sodium succinate group (2% sodium succinate + chow diet, n = 38). Each diet was provided for 16 weeks. Food was changed twice a week, and body weight was measured weekly during the 16 weeks of feeding. Carbon dioxide (CO2) was used as the euthanasia agent for mice.
Western blotting
The protein was isolated by lysing liver tissue using the WSE-7420 EzRIPA lysis kit (ATTO, Tokyo, Japan) containing a RIPA buffer, a protease inhibitor, and a phosphatase inhibitor. Equal amounts of loading proteins were loaded on an SDS/PAGE gel and further electro-transferred onto PVDF membranes before being blocked for 1 h with nonfat dry milk (5%) at room temperature. Primary antibodies were used to incubate overnight at 4°C and determine the levels of proteins. Primary antibodies included those specific for α-SMA (ab5694, abcam, Cambridge, UK) and GAPDH (GTX627408, GeneTex, Irvine, CA, USA). The membranes were incubated with secondary antibodies conjugated to HRP, and the Westsave Star Detection Reagent system (AbFrontier, Seoul, Korea) was used to detect protein bands. Loading control for western blotting was GAPDH.
Real- time PCR
Trizol reagent (Qiagen, Hilden, Germany) was used to isolate total RNA from mouse livers, which was then transcribed to cDNA using oligo dT (REF C110A, Promega, Madison, WI, USA) and hypescript RT master mix 601-710 (containing hyperscript reverse transcriptase, mixture dNTPs, reaction buffer, stabilizer, RNase inhibitor; provided by GeneAll, Singapore). Subsequently, the mRNA copy number of several markers was determined using real time PCR Quant studio 6 Flex (Life Technologies, New York, NY, USA) with SYBR Green. Primer information are listed with sequence 5'-3': α-SMA mouse (Forward: GACCCAGATTATGTTTGAGACC, Reverse: CAGAGTCCAGCACAATACCAG), GPR91 mouse (Forward: TCACTGTGGTGTTTGGCTACCT, Reverse: CCCTTATCATTGGCATAACTCTTTATC), GAPDH mouse (Forward: GCACAGTCAAGGCCGAGA, Reverse: GCCTTCTCCATGGTGGTGAA).
Triglycerides assay
The triglycerides assay kit (AM 157S-K, Asan Pharmaceutical, Seoul, Korea) uses an assay protocol where triglycerides are converted to free fatty acids and glycerol. Glycerol is oxidized to form a product that reacts with a probe to produce color. Absorbance was measured at 550 nm wavelength.
Alanine transaminase activity assay Kit
The alanine transaminase activity (ALT) assay kit (AM 102, Asan Pharmaceutical, Seoul, Korea) uses a protocol where ALT catalyzes an amino conversion reaction between alanine and α-ketoglutaric acid to produce pyruvic acid and glutamic acid. Pyruvic acid is detected in a reaction that converts a nearly colorless probe to a colored form. ALT can be calculated by measuring absorbance at 505 nm.
Immunohistochemistry
The liver tissues were used for H & E and Masson's trichrome staining after fixing with 10% formalin. Picro-Sirius Red, and α-SMA special staining were done to measure fibrosis using sodium succinate diet-fed mouse liver tissues and control liver tissue.
Human liver specimens
Normal human liver specimens and human livers with diseases such as alcohol cirrhosis, primary biliary cirrhosis, hepatocellular carcinoma, nonalcoholic steatohepatitis (NASH), and non-fatty NASH (n = 12 specimen each) were obtained through the Liver Tissue Cell Distribution System (Minneapolis, MN, USA), which was funded by NIH Contract #HHSN276201200017C.
Statistical analyses
All data were collected from at least three independent experiments. Results are expressed as means ± SD. Statistical significance between the experiments was determined by comparing two groups using the two-tailed Mann-Whitney U test. Statistical analyses were performed using Microsoft Excel (Microsoft, USA), and GraphPad Prism5, and P-values < 0.05 were deemed statistically significant.
Results
Changes in body weight and serum ALT and hepatic triglyceride levels in mice fed a sodium succinate diet-fed mice
To evaluate the effect of the sodium succinate diet on mice, we measured body weight and the levels of serum ALT weekly. The results showed that the diet containing 2% sodium succinate decreased body weight significantly compared to the control group from 6 weeks to 16 weeks of feeding (Fig. 1A). In addition, we measured ALT (Fig. 1B) and hepatic triglyceride content (Fig. 1C) in the serum at 4 weeks, 6 weeks, 8 weeks, 12 weeks, and 16 weeks to examine the effect of the sodium succinate diet on mouse liver damage. The results showed that the diet containing 2% sodium succinate increased ALT and hepatic triglyceride levels in the serum at 8 weeks, 12 weeks, and 16 weeks significantly, compared to control group. These finding suggest that a sodium succinate diet might reduce body weight within 6 weeks and increase serum ALT and hepatic triglyceride content in mice within 8 weeks of feeding.

Timeline of changes in body weight and serum alanine transaminase (ALT) and liver triglyceride levels in sodium succinate diet-fed mice. Statistical significance: *p < 0.05, **p < 0.01 and ***p < 0.001 versus the control group.
Expression of liver fibrosis proteins and the succinate receptor in sodium succinate diet-fed mice
To evaluate the effect of a sodium succinate diet on liver fibrosis proteins, we assessed the protein and mRNA expression of α-SMA in chronological order at 4, 8, and 16 weeks of feeding (Fig. 2). Although the sodium succinate diet-fed mice showed increased α-SMA protein expression at 8 and 16 weeks of feeding, no change was observed at 4 weeks of feeding compared to control (Fig. 2A). However, the protein and mRNA expression of GPR91 increased within 4 weeks of feeding compared to the control and then decreased gradually until 16 weeks of feeding. The protein and mRNA expression of the succinate receptor, GPR91, increased at an early stage (4 weeks) of sodium succinate diet feeding compared to α-SMA expression, which is a fibrogenic marker. These results suggest that a sodium succinate diet might increase the production of fibrotic proteins within 8 weeks.
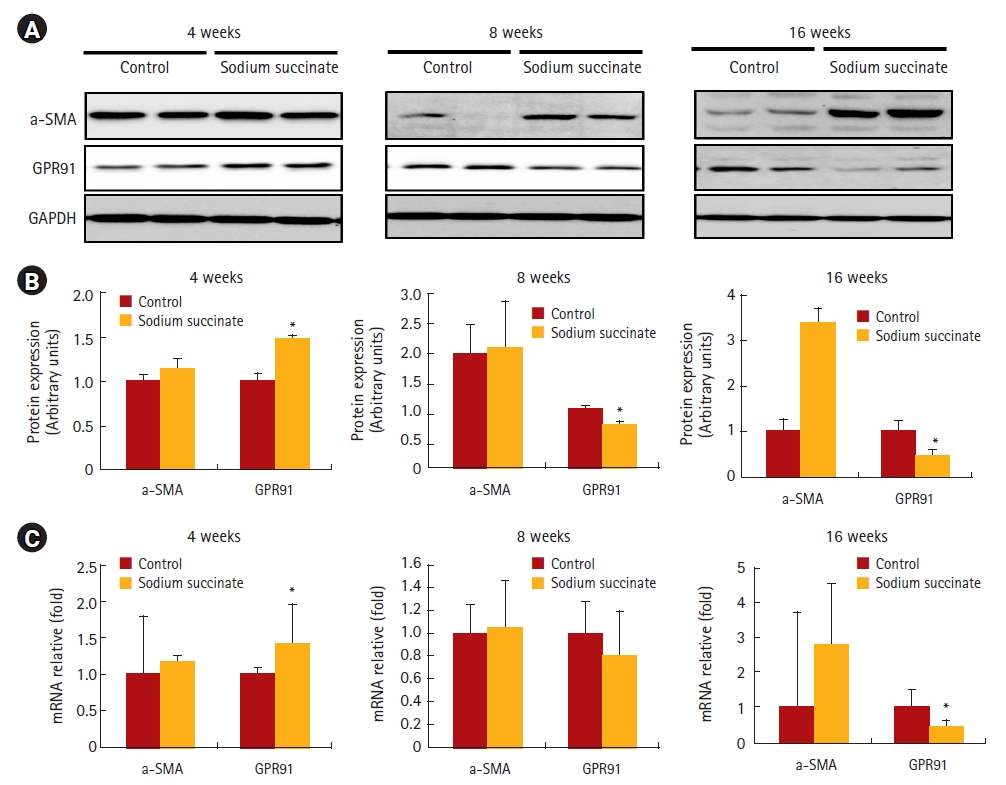
Chronological changes in protein and mRNA expression of alpha smooth muscle actin (α-SMA) and GPR91 in sodium succinate diet-fed mice. Liver specimens collected from the control group and 2% sodium succinate-containing diet group at 4 weeks, 8 weeks, and 16 weeks of feeding were used to analyze the expression of α-SMA and GPR91 by western blotting and RT-PCR. Densitometry analysis was performed, and data are presented as the mean ± S.E. of three independent experiments. Statistical significance: *p < 0.05, **p < 0.01 and ***p < 0.001 versus the control group.
(A) Western blot analysis of the protein expression of α-SMA and GPR91. (B) RT-PCR analysis of α-SMA and glucose transporter 1 mRNA expression.
Masson’s trichrome, Picro-Sirius Red, and α-SMA special staining of sodium succinate diet-fed mouse liver tissues
Masson’s trichrome staining showed that hepatic fibrosis increased significantly in sodium succinate diet-fed mice compared to the control group at 16 weeks of feeding (Fig. 3A), but not at 4 weeks or 8 weeks of feeding. In Picro-Sirius Red staining, sodium succinate group revealed more extensive collagen deposition at 16 weeks compared to control group (Fig. 3B). Immunostaining of a-SMA demonstrated that sodium succinate group showed increased amount of α -SMA positive cells compared to control group (Fig. 4).
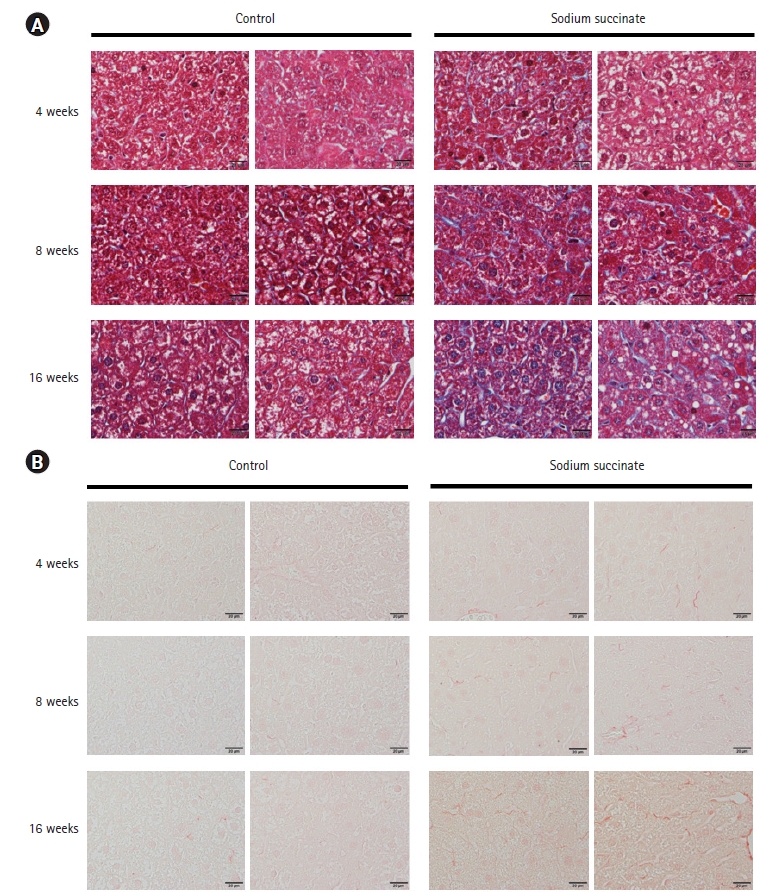
Representative images of liver tissues of sodium succinate diet-fed mice stained with Masson’s trichrome and Picro-Sirius staining. (A) Masson’s trichrome staining showed fibrosis in the liver tissues of sodium succinate diet-fed mice compared to control. (B) Picro-Sirius staining showed fibrosis in the liver tissues of sodium succinate diet-fed mice compared to control. All slides are magnified 400×.
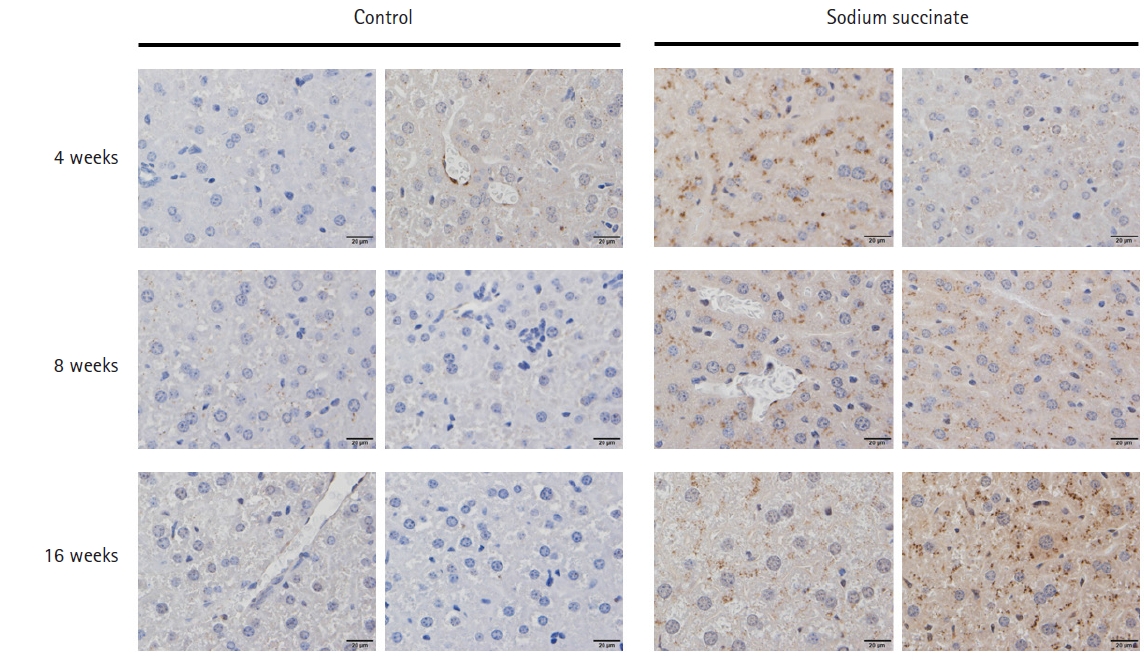
Representative images of liver tissues of sodium succinate diet-fed mice stained with a-SMA Immunostaining. Immunostaining of a-SMA demonstrated that sodium succinate group showed increased amount of α -SMA positive cells compared to control group. All slides are magnified 400×.
GPR91 expression was significantly decreased in non-fatty NASH human livers
We investigated the expression of GPR91 in normal, alcoholic cirrhosis, primary biliary cirrhosis, hepatocellular carcinoma, NASH, and non-fatty NASH human liver specimens. The results showed that GPR91 expression was significantly lower in non-fatty NASH and alcoholic cirrhosis liver specimens than normal samples, while no difference was noted in GPR91 expression between normal and NASH liver samples (p = 0.081) (Fig. 5).
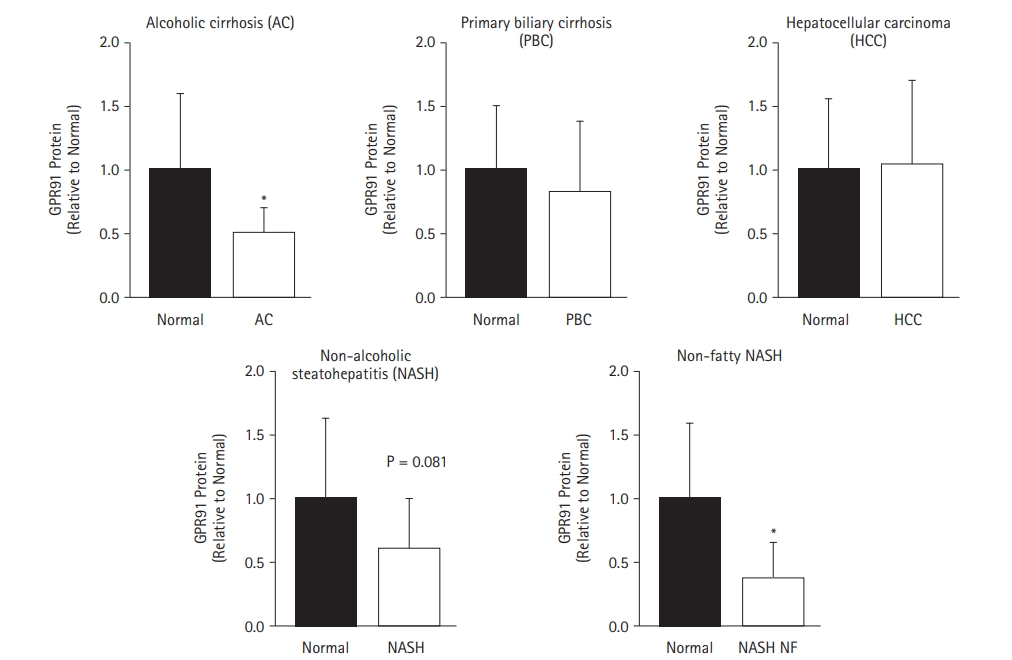
GPR91 protein expression in normal human liver specimens and human livers with various diseases. Western blot analysis of the expression of GPR91 in normal human liver and human livers with alcoholic cirrhosis, primary biliary cirrhosis, hepatocellular carcinoma, non-alcoholic steatohepatitis (NASH), and non-fatty NASH.
Discussion
In this study, we demonstrated that the sodium succinate diet administered for 16 weeks might induce the upregulation of α-SMA expression at 8 weeks of feeding (molecular changes) and then develop hepatic fibrosis (morphological changes) at 16 weeks of feeding in a time-dependent manner. We elucidate the possible role of GPR91 as an early fibrogenic marker, which was highly expressed at 4 weeks of feeding, even before the induction of molecular changes, such as the overexpression of α-SMA at 8 weeks of feeding.
Liver fibrosis involves the excessive accumulation of extracellular matrix proteins as the result of chronic liver injuries, such as chronic hepatitis B or C infection, alcohol, and free fatty acids in non-alcoholic fatty liver disease [7]. During the progression of hepatic fibrosis, HSCs play a major role [1].
Experimental mouse models of hepatic fibrosis can be induced by toxins (e.g., carbon tetrachloride and thioacetamide), diets (e.g., high fat diet and methionine- and choline-deficient diet), and surgery (e.g., bile duct ligation) [8]. In the present study, we focused on the effect of succinate within the liver to establish a new mouse model of liver fibrosis based on the sodium succinate diet. In our previous study, we found that daily administration of an intraperitoneal injection of 100 mg/kg succinate in C57BL/6J mice for four weeks could lead to increased α-SMA expression, but no histo-morphological changes, such as hepatic fibrosis, were seen [5]. The present study was designed to demonstrate that an oral succinate diet containing 2% sodium succinate instead of an intraperitoneal succinate injection, which was lethal over four weeks, may induce hepatic fibrosis through prolonged oral succinate feeding. In this study, we demonstrated for the first time that a sodium succinate diet leads to hepatic fibrosis changes in histology at 16 weeks of feeding, including the overexpression of the α-SMA protein and gene at 8 weeks of feeding. In addition, we clarified that the sodium succinate diet increased serum ALT and hepatic triglyceride levels in the mouse model. Our study suggests that the oral succinate diet mouse model could be a model of hepatic fibrosis or hepatic damage in future research.
The succinate receptor, GPR91, is expressed in quiescent rat HSCs, and the induction of ischemia in an isolated liver by interrupting portal flow leads to a marked increase in succinate release, thereby activating HSCs [9]. However, our previous study showed that the expression of GPR91 is significantly increased in activated HSCs, which is contrary to our findings in the present study [9]. Additionally, our previous study showed that GPR91 knockdown with small interfering RNA deactivates HSCs, demonstrating that succinate induces the activation of HSCs through GPR91 signaling [10].
Succinate accumulation and GPR91 signaling seems to be a key early step for cells to sense and respond to hyperglycemic conditions in the kidney, which leads to the activation of the intra-renal renin-angiotensin system [11,12]. Recently, another study showed that deletion of the GPR91 gene protected high-fat diet-fed mice from obesity only during the initial period; however, in later stages, body weight differences and weight curves were not different between GPR91 knockout and wild-type mice [13], suggesting GPR91 as an early sensor for dietary energy intake. Notably, during the development of liver fibrosis by the consumption of a sodium succinate diet, a marked overexpression of the GPR91 protein and gene in the liver occurred at 4 weeks of feeding, following which the expression of GPR91 declined rapidly from 8 weeks to 16 weeks. This might be explained by the compensatory response of GPR91 to sustained exposure to high concentration of succinate in long-term feeding.
Conclusion
Mice fed with sodium succinate showed increased expression of the α-SMA protein and gene in the liver at 8 weeks of feeding and demonstrated hepatic fibrosis histology at 16 weeks of feeding. However, the expression of the GPR91 protein and mRNA increased earlier at 4 weeks of feeding and then decreased rapidly from 8 weeks to 16 weeks. In addition, mice orally administered sodium succinate lost body weight and showed an increase in serum ALT and hepatic triglyceride levels compared to control mice. These results suggest that succinate could induce liver damage and fibrosis in the mouse model, and that GPR91 signaling might be an early marker or sensor of hepatic fibrosis development. Thus, GPR91 signaling could serve as a potential therapeutic target to prevent or delay the progression of liver fibrosis.
Notes
Conflict of interest
The authors declare no conflicts-of-interest related to this article.
Funding
This study has been worked with the support of a research grant of Kangwon National University in 2018 and NRF-2016R1C1B2011968 from Korea government.
Ethical approval
This research protocol for animal experimentation was accepted by the Institutional Animal Care and Use Committee of Kangwon University (KW-180103-1).